自閉症リスク因子が脳発生に与える影響を一網打尽に!
私達の遺伝情報をコードするゲノムの変異は疾患に結び付くことが知られている。
しかし、ノックアウトマウスの作成および解析は大変な労力が必要であり、多数の細胞種からなるヒトやマウスの体において、遺伝子の欠損がどの細胞に影響を与えるのかを調べるのは未だ簡単ではない。
そこで今回、シングルセルRNAseqとCRISPRを組み合わせた手法により、複数の自閉症リスク因子遺伝子が脳発生に与える影響を細胞種ごとに解析した論文を紹介する。
-----

自閉症は、対人関係の特異性やコミュニケーションの質的な障害などが見られる疾患(?)であり、これまでヒトを用いた研究により多数のリスク遺伝子が同定されている。
これらの遺伝子が、異なる特性を持つニューロンや、アストロサイト、オリゴデンドロサイトといった多数の細胞種からなる脳のどの細胞に影響を与えるのかを網羅的に調べるのは難しかった。
そこで筆者らはシングルセルRNAseqとCRISPRを組み合わせたPerturb-Seqを用いることで複数の遺伝子が神経発生に与える影響を解析することを試みた。

実験の流れは上の図のような感じ
ざっくりとは、
↓
Cas9を発現するマウスの胎児の脳にインジェクション(ウイルスが導入された細胞では標的遺伝子がノックアウトされることが期待される)
↓
ウイルスが導入された細胞をソートし、シングルセルRNAseq
↓
ウイルスにつけていたバーコードの情報から、それぞれの遺伝子を標的としたウイルスが導入された細胞を同定
↓
ノックアウトによって変動した遺伝子発現プロファイルの解析
という流れ。
(ちなみにメインFigureのデータだけで18セットのシングルセルRNAseqしているようだ。お金持ち!)
そういうわけで、35種類の遺伝子のノックアウトウイルスを導入した際にどの細胞種の遺伝子発現プロファイルが変動するかを検討した。
 興味深いのが赤丸で囲ったAnk2, Chd8, Gatad2bの3遺伝子である。
興味深いのが赤丸で囲ったAnk2, Chd8, Gatad2bの3遺伝子である。
これらの遺伝子は、これまで影響があるとは思われていなかったAnk2は抑制性ニューロン、Chd8とGatad2bはオリゴデンドロサイトの遺伝子発現プロファイルを変化させていることが分かる。
実際筆者らは、Chd8の(普通の)ノックアウトによってミエリンの形成が異常になる可能性を見出している。
(これはバイアスなしに変化を記述するPerturb-seqの強みがよく出ている結果のように思える)
最後はヒトの患者でみられた遺伝子発現変化が今回の系でも一部見られることを出している。(まあこれは解析の方法次第でどうにでも見せられそうだが)
------
コメント
・誰にでもできる、というほど簡単な手法ではなさそうだが、今後こういった網羅解析はより一般的になってくるのだろう。細胞種も比較的多い脳で、これまたリスク遺伝子も多い自閉症をモデルにこんなこともできる、というのを示した点では重要だと思った。
・こういう網羅解析は何段階ものクオリティコントロール、正規化、統計処理を重ねるため、こねくり回せば"ポジっぽい"Figureにできてしまうのではないかという懸念はある。やはり最終的には普通のノックアウトマウスなどの機能解析が必要だろう。
・今回は変化が見られる可能性が大きそうな候補因子だけに絞っていたが、もっと多数の遺伝子を対象とすることでこれまで脳発生に関係するとは思われていなかった遺伝子の寄与などを調べられると面白いかもしれない。
In vivo Perturb-Seq reveals neuronal and glial abnormalities associated with autism risk genes, Science, 2020
Xin Jin, Sean K. Simmons, Amy Guo, ..., Aviv Regev, Feng Zhang, Paola Arlotta
相分離を駆動する"ヌクレオソームコア"の構造変化
クロマチンは一般に緩い構造を取ると遺伝子発現が活性化しやすく、凝集した構造を取ると遺伝子発現が抑制されやすい。
このようにクロマチンの凝集度の制御は、遺伝子発現の制御にとても重要である。
これまで、クロマチンの凝集を制御する代表的な因子としてHP1(Heterochromatin protein 1)という因子が知られてきた。
しかしながら、HP1がどのような分子メカニズムで相分離を引き起こしているのかは不明である。

このとき興味深いことに、HP1の存在下ではH3とH3、H4とH4のように、コアヒストンの相互作用にも変化が起きていることが分かった。
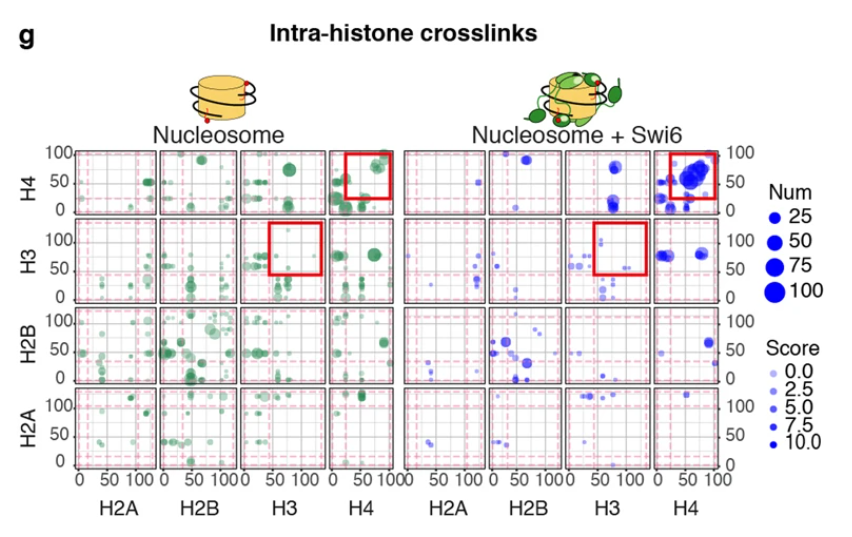
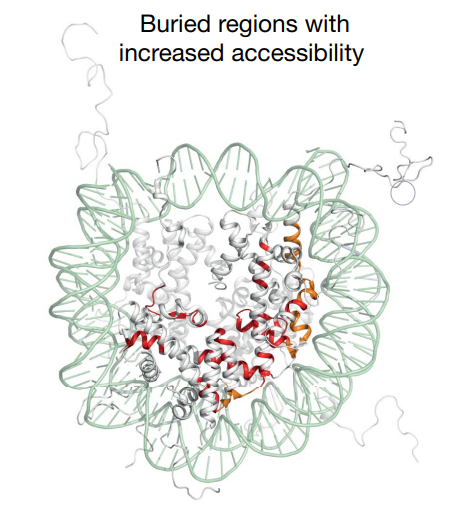
これは、HP1によってコアヌクレオソームの構造が変化している可能性を示唆する。
---
---
では、このヌクレオソームの構造変化はHP1の機能に重要なのだろうか?
また構造変化が起きにくいヌクレオソームを用いた場合、HP1による相分離能も低下していることが示された。
---
-----
コメント
・たしかにヌクレオソームの構造自体で凝集状態や相分離の状態が変わるというのは概念的に新しいかも。因果関係はヒストン変異体の実験しかないので、"ヌクレオソームの構造変化→凝集/相分離"がどれだけそれらしいかは疑問が残るが。
・細胞内でも同じようなことが起きているのか?というのはとても気になる。今回の場合、"ヌクレオソームの構造変化は起こさないHP1の変異体"が取れるとよいのだが、難しそう。
---
今回の論文
HP1 reshapes nucleosome core to promote phase separation of heterochromatin, Nature, 2019
S. Sanulli, M. J. Trnka, V. Dharmarajan, R. W. Tibble, B. D. Pascal, A. L. Burlingame, P. R. Griffin, J. D. Gross & G. J. Narlikar
真のオス化遺伝子の発見!?
今回珍しく日本人の方の論文を紹介させていただきます。関係者の方(に限らず)間違いなどございましたらご指摘いただけると幸いです。
----
性別をきちんと決定することは種の生存及び進化に非常に重要である。
これまで哺乳類のオス化を決定する遺伝子としてY染色体上のSryという遺伝子が同定されてきた。
Sryはその発見以来、単一のエクソンからなる遺伝子だと考えられてきた。
今回、Sryにこれまで未知であった第2エクソンが存在し、オス化に重要であることを示した論文を紹介する。

-----
まず、筆者らはRNAシーケンスを行うことで、Sry遺伝子座近傍で発現しているRNAを解析した。
このとき、通常であれば解析の段階でゴミとして切り捨てる「複数のゲノム領域に貼り付きうるリード」を捨てずに解析することで、これまで報告されてこなかったSry近傍の転写物を発見した。

(Sry遺伝子座は回文配列に挟まれ、平たく言うと面倒くさいゲノム領域に存在することから、マルチマップを許さないと見えてこなかったのだろう。いやー、マルチマップを許してみようと思う発想がすごい。)
筆者らはさらに転写開始点を特定可能なRNAシーケンス(CAGE-seq)や、タグをノックインしたマウスの作成を行うことで、このSry近傍の転写物はこれまでないとされてきたSryの第2エクソンであることを明らかにする。
筆者らはこれまでの単一エクソンからSryをSry-S(Single)、新しく同定した第2エクソンを含むSryをSry-T(Two)を名付けている。
Sry-TはSry-SのC端18アミノ酸が欠損し、新しく15アミノ酸を獲得したアミノ酸配列になっている。(以下の図を参照)


----
では、この第2エクソンを含むSry-Tは性決定に重要なのだろうか?
筆者らはSryの第2エクソンを削るようなマウスを作成し、解析を行った。
すると驚くべきことに、Sryの第2エクソンを削ったマウスでは染色体がXYであってもメスのようになることが分かった。

このことから、Sry-Tはオス化に必須の働きをすることが分かった。
---
さらに筆者らはSry-Tを発現するだけでオス化を誘導できるのか検証するため、XX染色体をもつマウスで生殖細胞特異的にSry-SおよびSry-Tを強制発現させ、解析を行った。
その結果、Sry-Sをヘテロで発現するマウスはメスのままであるのに対し、Sry-Tを発現するマウスはオス化することが分かった。
すなわち、Sry-Tの発現はマウスのオス化に十分であることが分かった。
-----
では、Sry-TはSry-Sと何が違うのだろうか?
興味深いことにSry-Sだけが持っているC末端のアミノ酸配列は、タンパク質分解メカニズムによって分解を受けやすい配列であることを見出した。
実際、蛍光タンパク質にSry-SのC末端の配列を付加すると安定性が(Sry-Tの末端配列を付加した場合に比べて)減少することを示している。
このことから、Sry-Tはタンパク質として安定であるためにオス化を誘導できる可能性が考えられる。
この可能性を検証するため、筆者らはSryの第2エクソンを削ったうえで、Sry-Sのタンパク質分解誘導配列に変異を加えたマウスを作成した。
Sryの第2エクソンを削ると染色体がXYでもメスになるが、さらにSry-Sのタンパク質分解誘導配列に変異を加えるとオス化することが分かった。
すなわち、Sry-TはSry-Sに比べタンパク質が安定である可能性が高く、その特性こそがオス化に重要である可能性が示唆された。
----
結果は大体以上で、今回筆者らによりこれまで見つかっていなかったSryの第2エクソンが見つかり、この第2エクソンを保持するタイプのSry-Tこそがオス化に重要であることがわかった。
どうやらこの第2エクソンはトランスポゾン由来の配列らしく、一度挿入が起こったのち、進化に有利な配列として残ったのだろう。
----
管理人コメント
・本当にとってもすごい!!RNA-seqのマッピングで重複を許して第2エクソンを発見し、マウスの劇的な表現型の発見にとどまらず、そのメカニズムにまで迫っていて震える。
・マッピングで重複を許したら見えてくるタンパク質とかまだまだあるのだろうか。本当にゴミだと思って捨てていたリードからすごいものが見つかったりして、、
今回紹介した論文
The mouse Sry locus harbors a cryptic exon that is essential for male sex determination
タンパク質ノックダウンで見えてきた新しい転写制御機構
遺伝子の発現がいかに制御されるか、という疑問は生物学において最も根源的な問題の一つである。
この重要性から、これまでにPolIIの活性を制御する因子が数多く報告されてきた。
この中でもNELF(negative elongation factor)という因子は1999年、東工大の山口先生、半田先生らによって、in vitroにおいてPolIIの転写を抑制する因子として細胞抽出液から同定された。
NELFは構造解析などからin vitroではPolIIの活性を抑えるとことが確からしいと思われてきたが、細胞内でNELFを欠損させるとむしろ遺伝子発現が下がるという報告もあり、統一的な見解が得られてこなかった。
このことから、その重要性にも関わらず、細胞内におけるNELFの一次的な機能は不明であった。
そこで今回、タンパク質ノックダウンによってNELFの一次的な機能について迫った論文を紹介する。

----
一般的なRNAのノックダウンやDNAを改変するノックアウトでは、多くの場合ノックダウン/アウトから解析するまでの時間が長い(>24時間)ためタンパク質の持つ一時的なな機能を解析することが難しかった。
そこで筆者らは薬剤依存的に目的のタンパク質を分解するAIDというシステムを導入した。
AID法は遺伝研の鐘巻先生らによって開発された手法であり、植物ホルモンのオーキシンを培地中に添加することでタグをつけたタンパク質を分解することができる。
筆者らはNELFにタグをつけることでオーキシン依存的にNELFを分解する系を立ち上げた。
以下の図のように、実際オーキシン添加30分でNELF-Cの量が大きく減少していることが分かる。すごい!!

というわけで、この素晴らしい系を用い、筆者らはNELFがなくなったときの転写の状態を新生RNAseq(PRO-seq)によって解析した。
すると、興味深いことに、NELFを分解するとTSS直下の+1ヌクレオソーム上でPolIIが停止する位置が少し後ろにずれることが分かった。
(メモ;おそらくSHL-6での停止からSHL-5/-2/-1への変化と思われるとのこと)
このことから、NELFは単にPol2を止めるのではなく、+1ヌクレオソーム上でPolIIが停止する位置を制御していることが明らかになった。

イメージとしてはこんな感じ。NELFがなくなるとPolIIがヌクレオソーム上でちょっとだけ進む。論文より引用。

---
さらに、筆者らはNELFがもつPol2の位置の制御以外の細胞内機能に迫った。
筆者らは、
・熱ストレスに対する素早いレスポンス
・RNAへのキャッピング反応
の2点について検討を行った
熱ストレス時には、停止していたPolIIがリリースすることで素早い転写を可能にしている可能性が報告されている。
そこで筆者らはNELFを分解させる条件で熱ストレスをかける実験を行った。
このとき意外なことに、転写量及びPolIIの停止位置はNELFがなくなっても大きな変化はなかった。このことから、NELFは熱ストレスに対するレスポンスには大きな寄与をしていないことが示唆された。
というわけで、つぎに筆者らはRNAへのキャッピング反応について検討した。
---
結果は以上。全体として、タンパク質を分解する系を用いることで、NELFのAcuteな機能に近づいた。
以下、Graphical abstract

-----
コメント
・RNAのノックダウンやノックアウトでは見えないこともあるというのは大事。こういうタンパク質分解の系はこれからも増えてくるのだろうか。
・今回はキャッピングとの関係を見ていたが、一般的にヌクレオソーム上で止まる位置による生物学的意義ってなんなんだろう。より奥で止まるようになっても発現量的にはそんなに変わらないんだろうか?
・ところでなんでNELFがないとpausingの場所が変わるのだろう?NELFがヌクレオソーム自体と相互作用していたりするのかな?
今回の論文
NELF Regulates a Promoter-Proximal Step Distinct from RNA Pol II Pause-Release, Molecular Cell, 2020
2020年ノーベル医学生理学賞;C型肝炎ウイルスの発見
2020年のノーベル医学生理学賞は「C型肝炎ウイルスの発見」でHarvey J. Alter, Michael Houghton, Charles M. Riceの3氏に授与されます。

Biostationでは、受賞対象となった研究についてまとめてみようと思います。内容は概ねノーベル財団の公式発表に基づいております。科学未来館のページも参考にしました。管理人はウイルスの研究者ではないので念のため。
----
肝炎は主にウイルス感染によって引き起こされます(アルコール摂取や環境要因、自己免疫疾患によっても引き起こされます)。
1940年代には既に感染性の肝炎には2つのタイプがあることが分かっていました。
一つは急性の肝炎であり、汚染された水や食べ物で感染し、一般的に長期的な影響は小さいとされます(今でいうところのA型肝炎)。
もう一つが、肝硬変や肝臓がんなどの慢性疾患の原因となりえるタイプの肝炎です(今でいうところのB型/C型肝炎)。

---
では、これらの慢性肝炎の原因は何でしょうか?
輸血による肝炎が問題になったのちの1960年代に、Baruch Samuel Blumbergらが、B型肝炎ウイルスを特定し診断検査と効果的なワクチンを開発しました(彼はこれらの業績により1976年にノーベル賞を受賞しています)。
今回ノーベル賞を受賞するHarvey J. Alterらは、この謎の肝炎を発症した患者の血液をチンパンジーに注射するとチンパンジーが肝炎を発症することから、この謎の肝炎は血液由来の物質によって感染することを突き止めました。
この肝炎は「非A非B肝炎」と名付けられ、さらなる研究によりこの感染の原因となる物質の実態はウイルスであろうこともわかってきました。このウイルスを以下しばらく、非A非B肝炎ウイルスとします。
---
今回ノーベル賞を受賞するMichael Houghtonらは、この非A非B肝炎ウイルスの単離に大きく貢献をしました。
網羅的な探索の結果、1つの陽性クローンが発見され、非A非B肝炎ウイルスはフラビウイルスファミリーに属するRNAウイルスであることが分かりました。
---
ここで残る重要な問題は、この非A非B肝炎ウイルスと思われるウイルスは本当に肝炎を引き起こすのか?という点です。
残る受賞者のCharles M. Riceらは、非A非B肝炎ウイルスのゲノムの特徴を解析するとともに、非A非B肝炎ウイルスに由来するRNAをチンパンジーの肝臓に注入すると、非A非B肝炎ウイルス肝炎の患者さんに類似した病理学的変化が見られることを見出しました。
すなわち、この非A非B肝炎ウイルスこそが、非A非B肝炎の原因であることが分かりました。
この非A非B肝炎ウイルスが、現在のC型肝炎ウイルスということになります。
---
3氏らの貢献をまとめると以下の図のようになります。
Harvey J. Alterらは非A非B肝炎は血中に含まれるウイルスによることを見出し、Michael HoughtonとCharles M. RiceらによりC型肝炎ウイルスの実体が明らかにされました。
これらの結果は、現在のC型肝炎の診断や治療薬の開発に繋がっており、非常に重要な発見であったと思われます。

---
新型コロナウイルスが流行している状況でのウイルス関連の受賞ということで、改めてウイルス研究の大事さを感じますね。
いつパンデミックが起きるか分かりませんし、地道に基礎研究を行うのも大事ですよね。頑張りましょう!!
---
受賞理由となった論文リスト(ノーベル財団ホームページより引用)
Alter HJ, Holland PV, Purcell RH, Lander JJ, Feinstone SM, Morrow AG, Schmidt PJ. Posttransfusion hepatitis after exclusion of commercial and hepatitis-B antigen-positive donors. Ann Intern Med. 1972; 77:691-699.
Feinstone SM, Kapikian AZ, Purcell RH, Alter HJ, Holland PV. Transfusion-associated hepatitis not due to viral hepatitis type A or B. N Engl J Med. 1975; 292:767-770.
Alter HJ, Holland PV, Morrow AG, Purcell RH, Feinstone SM, Moritsugu Y. Clinical and serological analysis of transfusion-associated hepatitis. Lancet. 1975; 2:838-841.
Alter HJ, Purcell RH, Holland PV, Popper H. Transmissible agent in non-A, non-B hepatitis. Lancet. 1978; 1:459-463.
Choo QL, Kuo G, Weiner AJ, Overby LR, Bradley DW, Houghton M. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science. 1989; 244:359-362.
Kuo G., Choo QL, Alter HJ, Gitnick GL, Redeker AG, Purcell RH, Miyamura T, Dienstag JL, Alter CE, Stevens CE, Tegtmeier GE, Bonino F, Colombo M, Lee WS, Kuo C., Berger K, Shuster JR, Overby LR, Bradley DW, Houghton M. An assay for circulating antibodies to a major etiologic virus of human non-A, non-B hepatitis. Science. 1989; 244:362-364.
Kolykhalov AA, Agapov EV, Blight KJ, Mihalik K, Feinstone SM, Rice CM. Transmission of hepatitis C by intrahepatic inoculation with transcribed RNA. Science. 1997; 277:570-574.
再生に伴う遺伝子発現のダイナミックな変動
再生の過程では一度分化した細胞が多分化能を獲得し、さらに元通りに再び分化するというダイナミックな分化能の変遷をたどる。
しかしながら、再生芽はたくさんの細胞種を含むため、再生過程において特定の細胞系譜での遺伝子発現がどのように変化するのかは全く分かっていなかった。
そこで今回は、分化能がダイナミックに変化する結合組織細胞をモデルに、再生過程における遺伝子発現変化に迫った論文を紹介する。

結合組織細胞は再生芽において最も多い細胞種であり、再生過程において骨と軟骨、腱、骨格周囲、真皮、間質性線維芽細胞など多数の細胞種を生み出すもととなる。
筆者らは遺伝学的に結合組織細胞をラベルするウーパールーパーを作成した(ちなみにPrrxでcreERT2を発現するライン、とてもよくValidationしている)。
次に、このラインを用いて再生過程おけるシングルセルRNAseqを行った。

その結果
・再生過程において、結合組織細胞は多様な遺伝子発現プロファイルを持つ状態から均一な細胞集団に変化すること(おそらくこれは分化した細胞が一度多分化能を獲得することに対応する)
・損傷直後に細胞外組織をリモデリングする遺伝子や細胞増殖を促す遺伝子の発現が上がってくること
などが明らかになった。
---
何もないところから組織を構築するという点で、再生は発生過程と似ている。そこで、筆者らは次に発生過程におけるシングルセルRNAseqを行うことで発生と再生の共通点と相違点を探索した。
この結果、
・損傷直後(損傷後10日目まで)はどの発生ステージの細胞とも異なる発現プロファイルになること
・再生過程の細胞は損傷後11日目ごろに、発生過程の細胞と似た発現プロファイルになること
が分かった。
すなわち、再生時には発生と異なる遺伝子発現の変化をたどって、発生時と似た遺伝子発現プロファイルに終着することが示唆された。
---
また、このシングルセルRNAseqの結果により、再生時の結合組織細胞はいくつかのサブ集団が存在することが分かった。
この結果、ラベルしたサブ集団は腕の根本側の再生に大きく寄与し、先端部の再生にはあまり寄与しないことが分かった。
すなわち、再生時に現れるサブ集団は異なる様式で再生に寄与している可能性が示唆された。
---
あとは再生芽から再び分化細胞が現れる過程の解析と系譜追跡をしているが力尽きたので割愛、、、
全体として、初めて特定の細胞系譜で再生過程における遺伝子発現を単一細胞レベルで明らかにしたことは新しい!
一応Graphical abstract

以下管理人コメント
・主流でもないモデル生物で系譜追跡できるラインを作ってシングルセルRNAseqと、かなり大変そうな実験をしていてすごい、!!
・やはりこの変化を制御する因子が分かると面白いですね!
・再生と発生で異なる遺伝子発現プログラムの詳細は何だろう?どうやって違いが生まれるのだろう?(再解析すればわかるが、、)
今回紹介した論文
Single-cell analysis uncovers convergence of cell identities during axolotl limb regeneration, Science, 2018
Bio-station, 2020, communicated by L.F.
細胞外タンパク質の品質管理
タンパク質の異常な凝集などは、神経変性疾患をはじめとして種々の疾患の原因となりうることから、タンパク質の品質は生体内で保たれる必要がある。
これまで細胞内でタンパク質の品質管理を担う因子は数多く同定されてきたが、細胞外タンパク質の品質がどのように担保されているかはあまり分かっていなかった。
今回線虫の系を用いて、細胞外タンパク質の品質管理を行うメカニズムに迫った論文を紹介する。

------
細胞外タンパク質の品質の異常の一つとして、タンパク質の凝集があげられる。
そこで筆者らはまず、細胞外タンパク質の凝集をモニターするような実験系を立ち上げた。
具体的には、細胞外に放出され、凝集しやすいことが知られていたLBP-2という因子に蛍光タグをつけた線虫を作成した。
次にこの線虫を用い、LBP-2の凝集が異常になるような変異体をRNAiスクリーニングで網羅的に探索した。
これらの因子はタンパク質の凝集を阻害するような機能があることが予想され、実際13遺伝子それぞれの欠損ではLBP-2の凝集が増加することをみている。
(ちなみにLBP-2だけでなくLYS-7というタンパク質の凝集でも同じようなことをみているので、これらの因子はLBP-2だけでなく多くのタンパク質の品質管理を担っていることが期待される。)
---
さらに筆者らはこれらの因子を過剰発現するだけで、凝集タンパク質が減少するかを検証した。
このために、ECRのうち、CLEC-1とLYS-3という因子に加え、機能が全く不明な遺伝子2つ(C36C5.5 and F56B6.6)を過剰発現するような線虫を作成し、解析を行った。
この結果、これら4つのECRそれぞれ単独の過剰発現によってLBP-2の凝集が減少することが分かった。
すなわち、ECRは過剰発現するだけで細胞外の凝集タンパク質を減らせることが分かった。
また興味深いことに、これら4つのECRはLBP-2と共局在し、ECRの少なくとも一つは免疫沈降でLBP-2との相互作用が検出されたことから、ECRは細胞外凝集タンパク質と直接相互作用して効果を発揮している可能性が示唆された。
----
筆者らはさらに、これまでタンパク質の品質管理と老化の関係が示唆されてきたことから、細胞外タンパク質の凝集と老化の関係に着目した。
結果をまとめると、LBP-2の凝集は老化に伴って増加すること、さらにECRの過剰発現により凝集は減少し、寿命が延びることを見出している。
ちなみに、免疫の活性化との関係も見ているが結果は省略。
以下がまとめ図。(News&Viewsから引用)

この研究では、細胞外タンパク質の凝集をモニターする系を確立し、細胞外タンパク質の品質管理を担いうる遺伝子を複数同定した。
------
コメント
・めっちゃ大事そうなのに細胞外タンパク質の品質管理メカニズムって意外と分かっていなかったのね。Questionがいかしてますね。
・ECRが細胞外タンパク質の品質管理を担うメカニズムは気になる。凝集したやつを分解に導くのか、そもそも凝集自体を防いでいるのかもちょっとよく分からない?あと凝集タンパクを認識する仕組みも知りたい。
・哺乳類にも今回見つかったECRのオルソログがあるらしい。哺乳類での機能も知りたいところ。
今回紹介した論文
Extracellular proteostasis prevents aggregation during pathogenic attack, Nature, 2020
Bio-station, 2020, communicated by L.F.